





La mayoría de las mediciones espectroscópicas en este trabajo se realizaron a temperaturas criogénicas de 4,5 K. La Figura 2 muestra todos los espectros experimentales relevantes (absorción, fluorescencia, excitación de fluorescencia polarizada y quema de agujeros) para los complejos LH1-RC de BChl. B-Contiene Sociedad Anónima. viridis De lado con los de Rhodobacter (Rba.) sphaeroides Y teorodovibrio Dinastía 970 (lujo. 970). Estos últimos complejos se presentan como referencia, al igual que los complejos que contienen BChl a con menos (Rba. esferoides) y más (lujo. 970) espectros de absorción desplazados al rojo, respectivamente. Los datos procesados para todos los complejos estudiados se muestran en la Figura 3.
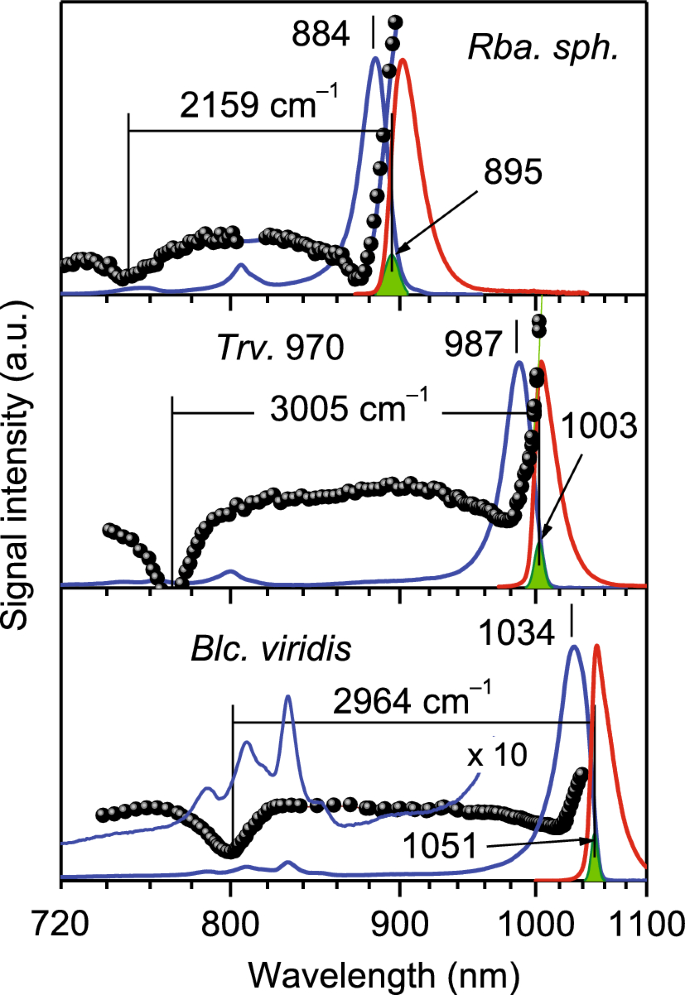
Espectros de absorción (azul), fluorescencia (rojo), excitación, contraste de fluorescencia (bolas negras) y combustión de agujeros (formas verdes) de complejos LH1-RC Sociedad Anónima. Viridis, lujo. 970 y Rba. esferoides Grabado a 4,5 K. Los espectros arbitrariamente normalizados se muestran en cuadros separados. área de la cola Sociedad Anónima. viridis El espectro se amplifica para mostrar la fracción de absorción de RC estructurada. Los números indican las posiciones espectrales de Q.y Las bandas de absorción de excitones y los máximos de los espectros de acción de quema de huecos (indicados por flechas) están en nanómetros. Las líneas horizontales miden los anchos de banda de los excitones en unidades de número de ondas.
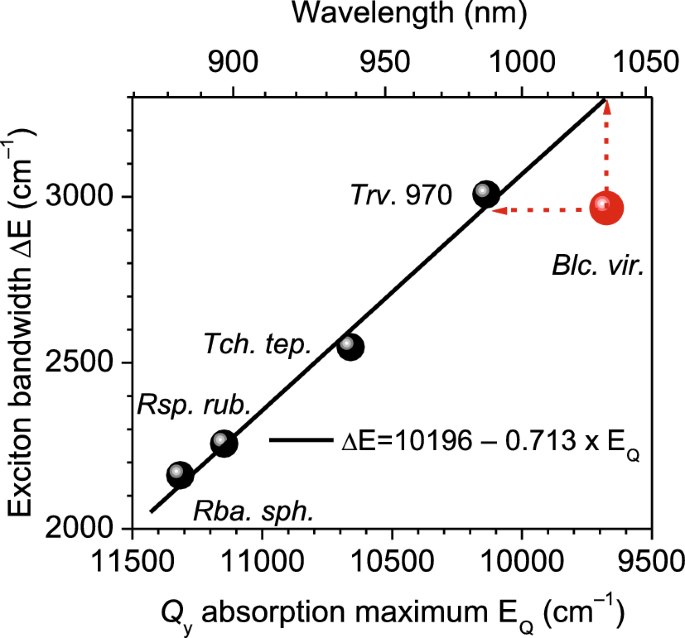
Anchos de banda de excitones ∆E con incertidumbre estimada de ±20 cm-1 Se determina a 4,5 K en función de Qy Energía de transición es. datos BChl a-Complejos que contienen bolas negras. El ajuste de regresión lineal a estos datos se muestra mediante la línea continua.
En los espectros de absorción de la Figura 2, los componentes espectrales LH1 y RC están bien separados entre sí. en Sociedad Anónima. viridisPor ejemplo, LH1 es responsable de que la banda asimétrica principal alcance un máximo de 1034 nm, mientras que un grupo de bandas más débiles que alcanzan un máximo de 788, 808, 819, 832 y 851 nm pueden unirse fuertemente a RC.18,19. La forma asimétrica del espectro LH1 es el resultado de múltiples estados de excitones superpuestos en el ensamblaje periódico de colorantes BChl estrechamente acoplados.20. El origen de la amplia protuberancia observada a 894 nm se analizará en detalle en una publicación separada. Aquí sólo señalamos que esto se debe al acoplamiento de los excitones LH1 a alta frecuencia (≥ 1500 cm-1) Paralizar B Vibraciones. Los espectros de los complejos de referencia muestran una estructura comparable, aunque con un desplazamiento hacia las posiciones azules de las respectivas bandas espectrales. También es digno de mención que, si bien los desplazamientos relativos de las bandas RC en BChl a– El BChl B– Contiene bacterias que corresponden aproximadamente a Qy La diferencia de energía de traslación entre a Y B Después de modificaciones en los pigmentos BChl, el desplazamiento relativo mucho mayor de las bandas LH1 no permite una interpretación tan directa.
Como se describe inicialmente en la referencia21, se pueden obtener datos importantes para evaluar el ancho de banda del excitón, denotado por ∆E, en complejos periódicos de LH mediante mediciones simultáneas de la excitación de la anisotropía de fluorescencia y los espectros de combustión de huecos. Luego, ∆E se define operativamente como la diferencia de energía entre la baja de alta energía del espectro de excitación de fluorescencia polarizada (que da lugar a estados de excitón en la parte superior de la banda de excitones) y el pico del espectro de acción de quema de huecos, que es un medida de la posición promedio del estado de excitón de menor energía22. Las líneas horizontales en la Figura 2 indican los anchos de banda de excitones específicos. Cabe señalar que los estados de excitón que definen los bordes de la banda de excitones son ópticamente muy débiles, restringidos por la simetría circular del complejo LH1 y la orientación casi plana de Qy Vectores de momento dipolar de colorantes.
Incluso un breve examen de los datos de la Figura 2 lleva a la importante conclusión cualitativa de que la intensa absorción del corrimiento al rojo observado en Sociedad Anónima. viridis No puede ser un resultado directo de un mayor acoplamiento de excitones. Esto se debe a que ∆E medido en Sociedad Anónima. viridis (2964 centímetros-1) es más estrecho en lugar de más amplio en comparación con incluir lujo. 970 (3005 centímetros-1) mientras que el espectro de absorción es de 47 nm (461 cm).-1) se desplaza más hacia el rojo.
∆E en función de Qy Frecuencia de transición de excitones, Espara todas las muestras analizadas en este trabajo se muestran en la Figura 3. Como se ve, se estableció una relación lineal exacta entre los datos de BChl a-Contiene complejos básicos descritos por la ecuación ∆E(E).s) = 10.196−0,713 × mis. Después de los datos vinculados Sociedad Anónima. viridis Se está desviando claramente de este camino. La coincidencia forzada requiere la consideración del ancho de banda de excitones mucho más amplio (∆E ≈ 3300 cm-1) o un valor mayor de Es Aproximadamente 10.143 centímetros-1 (~986 nm en la escala de longitud de onda). Estas dos opciones se indican en la Figura 3 mediante flechas discontinuas. La suposición anterior puede ser inmediatamente rechazada por las mediciones actuales que determinan el ancho de la banda de excitones. Sociedad Anónima. viridis. Sin embargo, esta última idea merece un estudio más profundo dada la observación de que la brecha de energía entre las bandas de absorción previstas (986 nm) y experimentales (1034 nm) es de 472 cm.-1 Está dentro del margen de error del 15%, que es lo mismo que el intervalo entre Qy Bandas de absorción para BChl a (a 771 nm) y BChl B (a 796 nm) disuelto en éter dietílico23.
La fuerte relación lineal observada entre ∆E y Es Dentro de complejos que contienen BChl a Sugiere fuertemente que existe un mecanismo dominante que impulsa el cambio de banda de absorción en los complejos LH1. Sorprendentemente, este mecanismo parece ser bastante resistente frente a muchas de las diferencias obvias demostradas en estudios recientes de microscopía crioelectrónica sobre complejos centrales bacterianos, como se informa en las referencias.6,7,8. Estas diferencias incluyen no sólo aspectos como los enlaces de hidrógeno de la proteína BChl y las conformaciones de BChl, discutidos anteriormente, sino que también se extienden a sitios de unión específicos para pigmentos proteicos, la presencia de iones metálicos y diferencias en el contenido de carotenoides. La bien conocida propiedad de los excitones no locales, conocida como su capacidad para mitigar el efecto de diversas perturbaciones estáticas y dinámicas (como se menciona en la ref.24) nos lleva a suponer que, en vista de las estructuras razonablemente comparables de todos los complejos estudiados, el comportamiento de los excitones en el hipotético complejo LH1 de Sociedad Anónima. viridisdonde cada BChl B Las moléculas son reemplazadas por BChl. a moléculas, se alineará estrechamente con la tendencia representada por la línea negra en la Figura 3.
Lo que hace que el conjunto de datos actual sea realmente especial es su capacidad para separar, por primera vez, las contribuciones de la sustitución de pigmentos (BChl). a a BChl b) y excitones en la respuesta espectral desplazada al rojo de cualquier sistema fotosintético original. Cuantitativamente, el ancho de la banda de excitones, representado por ∆E = 2964 cm-1lo que significa que la energía de acoplamiento más cercana del excitón es de aproximadamente 740 cm-1 El valor del desplazamiento del excitón es de aproximadamente 1480 cm.-1. Es importante destacar que este cambio es relativamente mayor que el cambio de energía del sitio estimado en 472 cm.-1. Desafortunadamente, lograr una separación más precisa de componentes de sintonización espectral condicionalmente distintos sigue siendo difícil de alcanzar en la fase experimental actual del estudio debido al componente de cambio de excitón no resuelto como se analiza en la ref.4.
Es posible que se haya observado una diferencia sistemática entre la posición de Es desde sy Los espectros de absorción de excitones de los complejos LH1 se han registrado a temperaturas criogénicas y ambientales, apareciendo estos últimos espectros en longitudes de onda más cortas. Para entender el origen de este fenómeno en el caso de Sociedad Anónima. viridisAdemás, se estudió la dependencia de la temperatura de sus espectros de absorción y excitación anisotrópica (Figura 4).
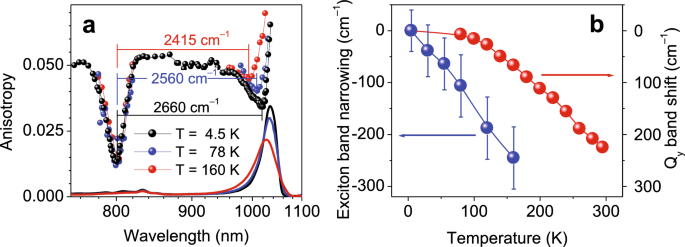
(a) Anisotropía de fluorescencia de excitación (datos dispersos) y espectros de absorción (línea continua) de complejos LH1-RC de Sociedad Anónima. viridis Grabado a temperaturas especificadas. (B) Dependencia de la temperatura de Qy Energía de transición de excitones (bolas rojas) y ancho de banda de excitones (bolas azules). Los datos se trazan en relación con los valores registrados a 4,5 K. Las líneas que conectan puntos de datos separados son para guiar la vista. Tenga en cuenta también que, a diferencia de la Figura 2, el ancho de banda del excitón se define aquí como la diferencia de energía entre las dos caídas en los espectros de anisotropía; consulte el texto para obtener más explicaciones.
Los datos de la Figura 4a muestran un estrechamiento del ancho de la banda del excitón con el aumento de la temperatura, en correlación aproximada con el desplazamiento hacia el azul de los espectros de absorción. Dado que la caída de anisotropía de alta energía mantiene su posición a todas las temperaturas, se espera que los cambios en la caída de anisotropía de baja energía y el espectro de absorción máxima se sigan con mayor o menos precisión. Como se muestra en la Figura 4b, existen grandes diferencias entre las tasas aparentes y las magnitudes de ∆E y E.s los cambios. Es probable que esto sea un artefacto resultante de la modificación asimétrica de la forma de la banda de absorción de excitones, que debido al frenado simétrico inducido térmicamente afecta principalmente al lado azul de Q.y banda, exagerando el cambio obvio ∆E. Observamos de paso que los excitones en BChl a-que contienen complejos LH1 revelan un comportamiento similardieciséis. La evaluación del borde de baja energía del ancho de banda del excitón mediante espectros de quema de agujeros, como se muestra en la Figura 2, probablemente habría reducido (si no eliminado) este aparente problema. Desafortunadamente, este enfoque más conveniente no es práctico aquí, porque las estrechas líneas de fonón cero en las que se basa la técnica existen en las proteínas fotosintéticas sólo a temperaturas bastante bajas, por debajo de aproximadamente 50 K.25,26.
En resumen, este estudio profundiza en los mecanismos detrás de la notable adaptabilidad de las bacterias fotosintéticas de color púrpura. Sociedad Anónima. viridis, que muestra una capacidad inusual para operar eficazmente en longitudes de onda muy largas del infrarrojo cercano. Este logro extraordinario, que va más allá de los límites de la actividad fotosintética, puede atribuirse a los efectos combinados de los mecanismos de conversión de energía del sitio y del excitón. Bacterias fotosintéticas dependientes de BChl a Su principal pigmento absorbente de luz prospera bajo luz infrarroja cercana más allá de aproximadamente 800 nanómetros, que se encuentra fuera del rango de absorción de la clorofila. Excitones asociados con BChl a Los pigmentos desempeñan un papel fundamental en la captación eficiente de luz en una amplia longitud de onda de 800 a 1000 nm (como se muestra en las Figuras 1 y 3). Sin embargo, como se muestra en este artículo, extender este rango más allá de 1000 nm requiere la participación de otro tinte (en la secuencia Chl a→ paralizar a→ paralizar B) siendo Q más rojoy Energía de transición. La limitación obvia en la capacidad de sintonización espectral de BChl aLos excitones pueden surgir de restricciones impuestas por la densidad de empaquetamiento del entorno proteico.
Si bien la naturaleza amplía naturalmente la gama de energía solar disponible para sustentar los ecosistemas, aprovechar esta energía para uso práctico representa un desafío tecnológico moderno crítico. Los conocimientos de este estudio sobre las funciones de los excitones y las transformaciones de energía del sitio en los sistemas fotosintéticos naturales tienen el potencial de inspirar futuras estrategias energéticas sostenibles y el desarrollo de dispositivos innovadores.

«Alborotador. Amante de la cerveza. Total aficionado al alcohol. Sutilmente encantador adicto a los zombis. Ninja de twitter de toda la vida».




More Stories
La Vía Láctea sobrevivió a una violenta batalla galáctica contra todo pronóstico científico
La NASA prueba helicópteros supersónicos para futuras misiones en Marte
La NASA descarta el impacto del asteroide 2024 YR4 contra la Luna en 2032